Parasitología
Parasitología
Introducción.
Giardia duodenalis (Sín: G. lamblia; G. intestinalis) es el nombre del protozoo flagelado del phylum Sarcomastigophora, subphylum Mastigophora, agente causal de la giardiasis (Monis et al., 2009), una parasitosis de intestino delgado proximal, cosmopolita, que puede manifestarse como un síndrome diarreico agudo, crónico o intermitente. También existe el estado de portador asintomático.
De acuerdo a la nueva sistémica en taxonomía, basada en datos bioquímicos, genéticos, y estructurales, Giardia pertenece al Phylum Metamonada, Subphylum Trichozoa.
La giardiasis es la protozoosis entérica más frecuente en el mundo. Desde el año 2004 fue incluida como una «enfermedad descuidada» (neglected disease) en la iniciativa de la OMS.
Transmisión. Epidemiología.
La enfermedad se contrae principalmente a través de alimentos y agua contaminados con materia fecal de hospederos infectados. Debe considerarse el potencial zoonótico derivado de la infección de Giardia. Afecta a diversos mamíferos, anfibios, reptiles y aves, de importancia el ganado lechero, animales de compañía y otros animales (Heyworth MF. 2016; Ryan & Cacciò. 2013; Ballweber et al., 2010). Estos animales representan reservorios potenciales importantes de Giardia; al respecto existen reportes de infección/enfermedad debidas asociadas a contacto con animales y sus genotipos (Murray et al., 2017).
Otros mecanismos que deben considerarse son: Contacto directo, este agente también puede ser transmitido por contacto sexual, exposición oral-anall, reportado sobre todo entre sujetos del sexo masculino, y a través de fomites. (Escobedo et al., 2014).
Los rotavirus, Cryptosporidium y Giardia han sido identificados como los principales agentes causales de episodios diarreicos en guarderías y asilos. La ubicuidad de Giardia es patente en ambientes urbanos, periurbanos y rurales en los que predominan higiene deficiente y hacinamiento.
Es importante considerar que existen a la venta diversos productos desinfectantes para eliminar bacterias, sin embargo, sólo algunos destruyen quistes y ninguno logra destruir huevos de helmintos (Soluciones desinfectantes – Información emitida por PROFECO en julio, 2007).
Se estima que alrededor de 280 millones personas/año en el mundo presentan la infección con diarrea aguda. Las infecciones asintomáticas son igualmente comunes (Ankarklev et al., 2010; Einarsson et al., 2016), observándose un mayor grado de riesgo en la población pediátrica.
En países dearrollados, la prevalencia relativa es de 0.2% – 29.2%, en contextos de brotes epidémicos y sin ellos: Desde 1960 la giardiasis se ha asociado a brotes epidémicos importantes en países altamente industrializados, por ingesta de agua contaminada y en guarderías. Actualmente, se reporta un aumento en el número de casos. Las prevalencias más altas se presentan en regiones tropicales y subtropicales de países en desarrollo. En México, la prevalencia e incidencia más altas se encuentran entre lactantes, preescolares y escolares.
Se ha demostrado la contaminación de fuentes de agua, ya sea redes de agua potable, de superficie o recreativas con microorganismos, entre ellos ooquistes de Cryptosporidium y quistes de Giardia, muy resistentes, lo cual significa un importante mecanismo de transmisión para humanos y animales.
– Pueden acceder a información epidemiológica en el Boletín Epidemiológico del Sistema Nacional de Vigilancia Epidemiológica Sistema Único de Información.
Los estudios moleculares han identificado 8 grupos genéticos o ensamblajes (A – H). (Heyworth MF. 2016; Benere et al. 2012; Asher et al., 2012; Bonhomme et al. 2011).
Los subgrupos incluidos en los ensambles A y B tienen mayor potencial zoonótico y se encuentran tanto en humanos como en animales. En México se han identificado con mayor frecuencia los ensamblajes AI y AII de perros. (Ryan & Cacciò. 2013; Asher et al., 2012; Cacciòa et al., 2010; Monis et al., 2009; Eligio-García et al., 2008; Ponce-Macotela et al., 2005).
Ensambles de Giardia |
Hospederos |
A |
Humanos, perros, gatos, ganado, alpacas, venados, hurones, cerdos, castores, chinchillas, jaguares, caballos, marsupiales, ovejas, cabras, buey almizclero, primates no humanos, cetáceo(s), focas, león de mar australiano, alce, reno, pollos, gaviotas |
B |
Humanos, ganado, perros, gacelas, venados, caballos, castores, rata almizclera, chinchillas, hurones, conejos, hutia cubana, marsupiales, cobayos, damanes, primates no humanos, pollos, ovejas, focas, cerdos, león de mar australiano, avestruces, delfines, marsopa, gaviotas |
C |
Perros, canguros, ganado, cerdos, cetáceos |
D |
Perros, chinchillas, canguros, ganado, cetáceos, zorros |
E |
Ganado, ovejas, cerdos, alpacas, cabras, caballos yaks, zorros, venados, gatos |
F |
Gatos, cetáceos, cerdos |
G |
Ratas, ratones |
H |
Foca gris, gaviotas |
Modificado de: Heyworth MF (2016) |
|
Morfología.
Giardia es un protozoo no invasivo, microaerofílico. Reside y se multiplica por división binaria en la superficie de las primeras porciones del intestino delgado, a un pH ligeramente alcalino que favorece su desarrollo y existe evidencia genética y epidemiológica sobre su capacidad de recombinación sexual (Monis et al., 2009; Cacciòa et al., 2010).
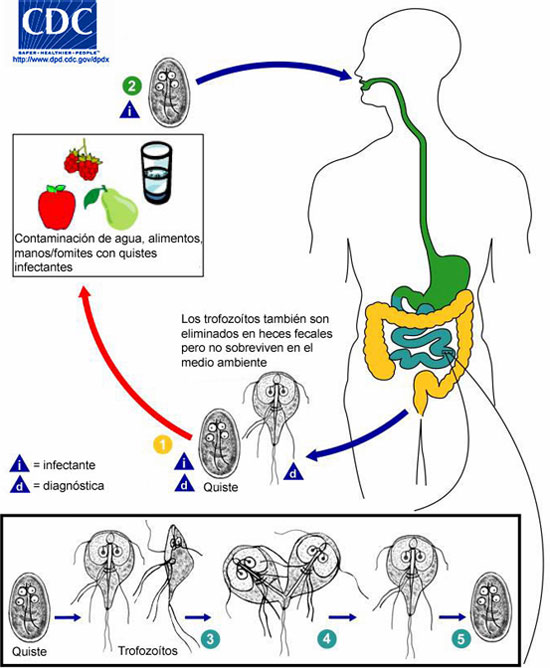
Presenta dos formas: trofozoíto y quiste con un ciclo biológico sencillo. El trofozoíto prolifera (se multiplica) y el quiste es la forma infectante
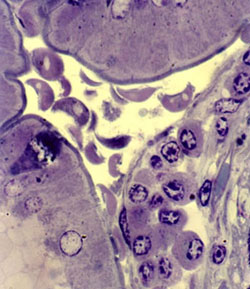
Trofozoíto G. duodenalis. S.J. Upton, Kansas University
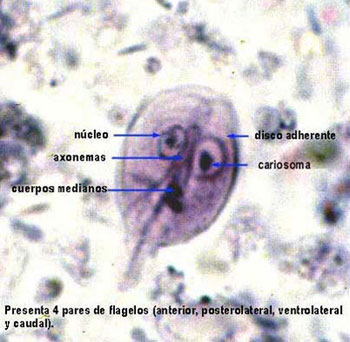
Los trofozoítos, formas vegetativas, miden 10 – 12 µm de longitud, son piriformes, con superficie dorsal convexa y ventral cóncava. Sus movimientos en espiral dan la impresión de «una hoja de árbol que cae». Las estructuras internas que pueden apreciarse son: dos núcleos con endosoma, cuerpos medianos en número variable*, disco adhesivo, ventral, con estructura cóncava, rígida, en espiral, de ~9 µm de diámetro, compuesto por microtúbulos y proteínas asociadas* ubicado en la mitad anterior ventral, con capacidad contráctil, y un paquete de axonemas con cuerpos basales en posición anterior con respecto a los núcleos, del cual derivan 4 pares de flagelos* (par anterior, dos pares laterales y par posterior) con el típico arreglo de microtúbulos 9+2. Carecen de mitocondrias y peroxisomas, y presentan mitosomas minúsculos <2 µm y nucleolo. El retículo endoplásmico rugoso y Golgi son aparentes durante la secreción de componentes requerida para el enquistamiento.
En ambientes con restricción de oxígeno, la mitocondria ha sufrido modificaciones importantes en contenido y función, dando lugar a «organelos relacionados con la mitocondria» (mitochondrion-related organelles – MROs), entre ellos los mitosomas, que no contienen ningún genoma y han perdido la capacidad de generar energía, pero retienen ciertas características de las mitocondrias, entre ellas la síntesis de grupos hierro-sulfuro (Einarsson et al., 2016) y los hidrogenosomas. Cuentan con estos organelos múltiples microorganismos parasitarios y de vida libre. Cabe mencionar a protistas anaeróbicos o microaerofílicos, como Entamoeba histolytica, Giardia intestinalis, Trichomonas vaginalis, Cryptosporidium parvum, Blastocystis hominis. El mitosoma de Giardia intestinalis presenta una importante reducción de proteínas y retiene una función mitocondrial: la biosíntesis de Fe/S
(Heinza E & Lithgow T. 2012; Makiuchi T & Nozaki T. 2013; Nyindodo-Ogari et al., 2014).
El citoesqueleto, necesario para la adhesión, desprendimiento y movimiento, tráfico intracelular, está compuesto principalmente por los 8 axonemas y sus cuerpos basales, los cuerpos medianos, el funis (capas de microtúbulos asociados a los axonemas de los flagelos caudales) y el disco adhesivo, en cuya composición se encuentran: alfa y beta tubulinas, giardinas, entre otras proteínas.
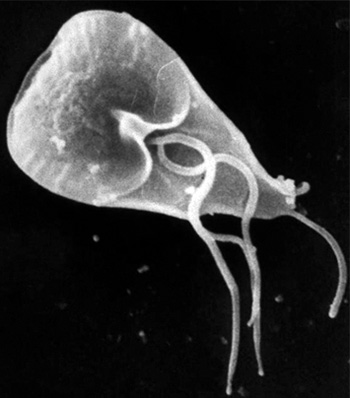
Giardia intestinalis. Trofozoítos. Microscopía de barrido: Se observa disco ventral y flagelos. Imagen: CDC/Janice Haney Carr.
Los quistes, formas de resistencia, infectantes, ovales, miden entre 11-14 µm de longitud y contienen 2 a 4 núcleos, de acuerdo a su madurez, y estructuras residuales de la forma vegetativa (axonemas, restos de disco adhesivo y cuerpos medianos). La resistente pared quística está formada por una capa filamentosa externa y una capa membranosa interna. Su grosor es de 0.3 – 05 µm. El principal carbohidrato del componente glicoprotéico externo es N-acetilgalactosamina (GalNAc).
Después de la ingestión, la exposición al ácido gástrico induce la activación del quiste en reposo. En respuesta al pH alcalino, las proteasas del intestino y señalizaciones propias del parásito, emerge una célula que se divide 2 veces sin replicación del DNA, produciendo eventualmente cuatro trofozoitos.
El enquistamiento inicia hacia la parte inferior del intestino delgado. La célula parasitaria detecta cambios en su hábitat y reponde formando una pared quística que le permite sobrevivir fuera del hospedero y hasta en agua fría cuando los quistes son eliminados con la materia fecal. Los quistes son transmitidos a otro hospedero, directamente, o a través de vehículos como agua y alimentos. Se estima que 10 – 100 quistes son suficientes como dosis infectiva (Ankarklev et al., 2010; Einarsson et al., 2016).
Ciclo Biológico
Patogenia.
La giardiasis es una enfermedad diarreica multifactorial y diversos mecanismos están involucrados. Existe la evidencia de trabajos experimentales in vivo, in vitro y estudios sobre la infección en el humano, proceso en el que se encuentran involucrados aspectos inmunológicos y funcionales de hospedero y parásito (Cotton et al. 2011; Ortega-Pierres et al. 2009; Gerbaba et al., 2015; Einarsson et al., 2016):
– Acortamiento difuso de microvellosidades e inhibición de las enzimas del borde de cepillo, con adhesión de los parásitos y células CD8+; hipersecreción de aniones (Cl-, con malabsorción de glucosa, sodio y agua) e hipermotilidad intestinal (en conjunto con el aumento en la permeabilidad), e:
– Insuficiencia y disminución de la actividad de algunas enzimas digestivas (ej. lactasa, proteasas, lipasas).
– El factor mécanico: adhesión mediante el disco ventral, movimiento flagelar; Alteraciones en las funciones de la barrera intestinal a nivel de los complejos de unión entre células epiteliales – F-actina, zonula occludens -ZO-1, claudin-1, y a-actinina, (alteraciones en las uniones estrechas).
– Inducción de quimiocinas en células intestinales epiteliales de ratón (IECs), lo que atrae células inmunes, como células cebadas y dendríticas.
– Inducción de apoptosis. Aumento en la apoptosis de los enterocitos; y aumento en el índice mitótico: rápido recambio celular (con células inmaduras). El tiempo de recambio celular normal es 3 – 6 días
– Deficiencia de arginina en in IECs, lo que da lugar a menor NO, detención del ciclo celular y apoptosis.
– Hiperplasia de las criptas y aumento en la secreción de moco.
– Cambios en la composición de la microbiota intestinal, disbiosis (Fink & Singer. 2017)
– Las diferencias genéticas entre grupos de Giardia
– Variación antigénica – proteínas de variación de superficie (VPSs).
– Edad, y estado nutricional del hospedero.
– Competencia por productos del hospedero: sales biliares, colesterol y fosfolípidos; Giardia no sintetiza de novo fosfolípidos y esteroles celulares (Cotton et al. 2011).
– Hiperplasia de las células caliciformes, lo que produce espacios celulares en la barrera epitelial (Ponce-Macotela et al., 2008).
– Existen estudios sobre giardiasis invasiva. Se requieren estudios para determinar el papel de los trofozoítos intraepiteliales de Giardia. (Reynoso-Robles et al., 2015; Martinez-Gordillo et al., 2014).
Patología.
Alteraciones en el borde «en cepillo» de las microvellosidades intestinales.
Atrofia o acortamiento de vellosidades.
Hiperplasia de las criptas.
Incremento en la permeabilidad celular.
Inflamación de la mucosa.
Sobrecrecimiento de poblaciones bacterianas.
Espectro clínico.
El período de incubación es de 1 – 2 semanas. Los cuadros clínicos oscilan entre el estado de portador asintomático y las enfermedades aguda y crónica.
Un gran porcentaje de personas presenta infecciones asintomáticas, con malabsorción intestinal imperceptible, que se resuelven espontáneamente.
Entre las manifestaciones de la enfermedad aguda se encuentran: diarrea acuosa o pastosa, esteatorrea (evacuaciones grasosas, generalmente explosivas y fétidas), dolor epigástrico postprandial, anorexia, distensión abdominal, flatulencia y ocasionalmente, cefalea, febrícula, manifestaciones alérgicas (artralgias, mialgias, urticaria); la enfermedad puede resolverse en unas semanas, aún sin tratamiento, pero la presencia de parasitosis crónica es un hallazgo cada vez más frecuente, actualmente, a nivel mundial, asociada con síndrome de intestino irritable, alergias alimentarias y urticaria (Einarsson et al., 2016), asi como con diarrea recurrente, esteatorrea, evidencia bioquímica de malabsorción de grasas, lactosa y otros disacáridos, vitamina A y vitamina B12, disminución de peso, deficiencias en el crecimiento y cognitivas en la población infantil (Certad et al., 2017; Bartelt & Platts-Mills. 2016).
Diagnóstico diferencial: Deben contemplarse rotavirus,adenovirus, Campylobacter, E. histolytica, Cryptosporidium, Escherichia colienteropatógena, Strongyloides stercoralis, enfermedad celiaca, úlcera duodenal.
Diagnóstico.
– Antecedentes epidemiológicos y cuadro clínico.
– Observación microscópica de trofozoítos (en materia fecal acuosa – mediante el examen directo en fresco, con solución salina y lugol) y quistes (en materia fecal sólida o semisólida – se utilizan exámenes coproparasitoscópicos de concentración por flotación), estudios de baja sensibilidad y alta especificidad. Si resultan negativos, se opta por ELISA para captura de coproantígenos. (Secretaría de Salud, CENETEC, 2012); varios estudios han reportado especificidades de 87–100% y sensibilidades de 63–100% y algunos falsos negativos, corroborados en mediante Técnicas basadas en la reacción en cadena de la polimerasa (PCR) (Koehler et al., 2014).
– Endoscopía con examen de contenido duodenal y biopsia intestinal son procedimientos invasivos, poco utilizados.
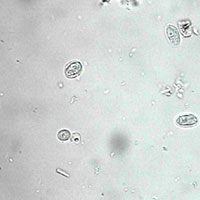
Examen directo. Solución salina. Quistes de Giardia. J.C.Fox, Oklahoma State University
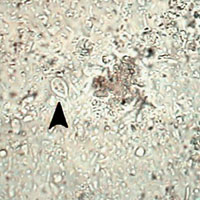
Examen directo. Solución salina. Trofozoitos de Giardia. J.C.Fox, Oklahoma State University
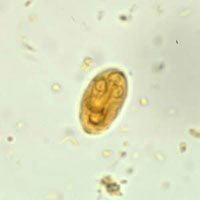
Quiste de Giardia. Tinción: lugol. DPDx/CDC.